Highlights
- Sex differences in longevity among model organisms are largely condition dependent
- Humans are unusual: female survival advantage is ubiquitous and persists throughout life
- Women live longer but suffer more physical limitations than men in later life
- Mechanisms underlying sex differences in longevity are poorly known
Abstract
Sex differences in longevity can provide insights into novel mechanisms of aging, yet they have been little studied. Surprisingly, sex-specific longevity patterns are best known in wild animals. Evolutionary hypotheses accounting for longevity patterns in natural populations include differential vulnerability to environmental hazards, differential intensity of sexual selection, and distinct patterns of parental care. Mechanistic hypotheses focus on hormones, asymmetric inheritance of sex chromosomes and mitochondria. Virtually all intensively studied species show conditional sex differences in longevity. Humans are the only species in which one sex is known to have a ubiquitous survival advantage. Paradoxically, although women live longer, they suffer greater morbidity particularly late in life. This mortality-morbidity paradox may be a consequence of greater connective tissue responsiveness to sex hormones in women. Human females’ longevity advantage may result from hormonal influences on inflammatory and immunological responses, or greater resistance to oxidative damage; current support for these mechanisms is weak.
Main Text
Introduction
The vast majority of animal species have two sexes, and those sexes often differ in many aspects of their biology. Most obviously, males range from a tiny fraction of the size of females to considerably larger and live considerably shorter to substantially longer lives (Austad, 2006; Finch, 1990). Sex differences in longevity can potentially be exploited to help understand mechanisms underlying variation in longevity within a species. Yet, these differences remain little studied despite considerable variation among different genotypes of commonly used laboratory species such as C. elegans (McCulloch and Gems, 2003), Drosophila melanogaster (Malick and Kidwell, 1966), and Mus musculus (Austad, 2011).
Surprisingly, much more has been reported about sex differences in longevity among wild populations than among captive populations because numerous long-term field studies have tried to understand the evolutionary forces underlying sex differences in behavior and a host of life history traits such as rates of development, mating systems, and reproductive patterns. Thus, we know that female short-finned pilot whales live nearly twice as long as males (Kasuya and Marsh, 1984) and that female African lions, red deer, black-tailed prairie dogs, numerous monkeys, and apes are also longer-lived than males, although these differences are not as extreme as in the pilot whales (Bronikowski et al., 2011; Clutton-Brock, 2009; Clutton-Brock and Isvaran, 2007). The reputed general pattern among mammals in nature is for females to be longer-lived, although this is known for certain in relatively few species. We know there are a number of mammals such as Japanese macaques, savannah baboons, and bannertail kangaroo rats where there is virtually no sex difference (Clutton-Brock and Isvaran, 2007; Waser and Jones, 1991). Rarely, male mammals even appear to be the longer-lived sex (see below).
The putative mammalian pattern of greater female longevity has been attributed to a variety of factors. George Williams (1957) hypothesized on theoretical grounds that sex differences in longevity likely reflected evolutionary histories of differences in vulnerability or exposure to environmental hazards due to differences in body size, foraging patterns, or other risky behaviors, which lead to sex differences in the rate of aging itself. Observations of sex-biased survival patterns in various species are consistent with this idea; however, the underlying theory predicting that differences in extrinsic mortality by itself leads to predictable evolved patterns of aging is now seen as incomplete at best (Abrams, 1993; Moorad and Promislow, 2010). Alternatively, sexual selection has been hypothesized to play a major role in longevity differences between the sexes as sexually selected traits are expected to exhibit an evolutionary fitness cost (Promislow, 1992; Promislow et al., 1992). Thus, the prevalence of polygynous mating systems in mammals concomitant with greater intrasexual selection among males could explain this general pattern if it truly exists (Trivers, 1972). A related, more narrowly defined, idea is that patterns of parental care affect sex differences in longevity. In support of this idea, females are the longer-lived sex in many primates, but in the few species such as owl and titi monkeys with extensive male parental care, males have a slight longevity advantage (Allman et al., 1998).
There are two additional hypotheses based on asymmetric inheritance. One of these is the heterogametic sex (or unguarded X) hypothesis, which posits that males are the shorter-lived sex because they have only a single X chromosome (Trivers, 1985). Specifically, the logic of this hypothesis is that because of females’ two X chromosome copies, the phenotypic effect of sex-linked deleterious alleles can be masked by normal alleles on the other X, whereas males have no second X chromosome to compensate for a single bad allele. A second hypothesis also based on asymmetric inheritance is that because mitochondria are only maternally inherited, they will be under selection for optimized compatibility only with the female nuclear genome. Therefore, any sexually antagonistic male genes would shorten male life (Tower, 2006).
The sexual selection, parental care, or the heterogametic sex hypotheses would each predict a different sex-longevity pattern in birds. Social monogamy is by far the most prevalent mating system in birds (Promislow et al., 1992; Trivers, 1972). Thus, intrasexual competition is expected to be reduced, and extensive male parental care is common, so predictions from sexual selection or parental care theories are that there would be little longevity difference between bird sexes. In addition, female birds are the heterogametic sex, so if the heterogametic sex hypothesis is valid, males would be predicted to be the longer-lived sex in birds (Austad, 2011). In fact, there is little reported longevity difference between the sexes in most bird species studied, but there are a range of species such as the Barnacle Goose, Acorn Woodpecker, and Arabian Babbler in which males are substantially longer-lived than females in the wild (Clutton-Brock and Isvaran, 2007). This is seen in both socially monogamous and socially polygynous species, consistent with the heterogametic sex hypothesis, but not with the sexual selection hypothesis.
Most of the studies adduced above are from wild populations. However, longevity differences in wild populations, while evolutionarily informative, may be due to a variety of factors unrelated to sex differences in intrinsic aging rate—the feature of most interest to laboratory and biomedically oriented biologists. Wild populations could exhibit sex differences in longevity simply because of differential risk-taking behavior, foraging patterns, or mortality due to direct sexual competition. For instance, among birds and mammals, one sex tends to be the dominant disperser—the one that leaves the natal territory to find a new territory, often at some distance. Dispersal, which involves traveling through unknown terrain, searching for new, available territory, is hazardous. In one study, water voles were found to have nearly 100 times the mortality rate during dispersal as when not dispersing (Leuze, 1980). Interestingly, one of the most sex-biased survival differences known among wild mammal populations is seen in Brandt’s bat (Myotis brandti), the longest-lived mammal species for its body size. Weighing only 7–8 g, M. brandti has been reported to live at least 41 years in the wild. All of the 67 individuals of this species known to have survived longer than 20 years have been males (Podlutsky et al., 2005). This could indicate that males of this species are physiologically protected against the ravages of senescence much better than females are. However, it is also possible that the extreme foraging behavior of lactating female bats that must consume up to 50% of their body weight in insect prey daily exposes them to greater risks of predation or energetic stress than males. Thus, the longer life of males could be due to differential exposure to environmental hazards rather than intrinsic aging processes. Differences in foraging or risk-taking behaviors are not the only factors complicating longevity differences between sexes. There is also reproduction itself. Reproduction, for instance, is known to be life-shortening in females of a number of species (de Heij et al., 2006; Liker and Székely, 2005). However, it can also be costly in males, particularly if one includes the cost of male competition (Iliadi et al., 2009; Nussey et al., 2009). Therefore, it is always wise to question whether sex differences in longevity observed in wild population reflect intrinsic differences in the rate of physiological senescence or not. Thus, while studies of sex differences in wild populations can provide important insights into the ultimate causes of aging, they are less useful for understanding the mechanistic underpinnings of sex differences in the intrinsic rate of aging.
If the goal is to understand sex differences in the cellular and molecular physiology of aging, studies of protected populations such as those found as pets or in zoos or research facilities are likely to be more informative. Easy generalizations about certain model species are often found in the literature. For instance, one can find statements that female flies are longer-lived or C. elegans hermaphrodites are longer-lived. Similar statements exist concerning sex differences in rodent longevity, sometimes favoring females, other times males. As we show below, such generalizations are seldom justified. Therefore, the rest of this review will focus on what is known about longevity differences in protected populations, focusing mainly on the most thoroughly known and studied laboratory species.
Worms
Mechanistically, longevity is arguably better understood in the nematode, C. elegans, than in any other animal species. Worms have two sexes, males and hermaphrodites. However, virtually everything we know about worm longevity has been from investigations of hermaphrodites only. In the standard N2 laboratory strain of C. elegans, which sex is longer-lived depends on culture conditions. Isolated males are 10%–20% longer-lived than hermaphrodites, yet when housed in groups, either mixed-sex or all male groups, males constantly attempt to mate, and the act of mating and mating attempts are such that there is no significant sex difference in longevity (Gems and Riddle, 2000). These results are reminiscent of those seen in Mediterranean fruit flies (Ceratitis capitata) in which sex differences in longevity vary depending on housing conditions, diet, and stress level (Carey and Liedo, 1995).
To determine whether this result was a quirk of the N2 strain or more general of the species as a whole, McCulloch and Gems (2003) reared 12 wild isolates that were highly divergent in a variety of traits. In two of these, the males appeared sickly through life and were exceptionally short-lived. Excluding these two, in 8 of the 10 other strains males were significantly longer-lived, and in the other two there were no significant differences. The mean ratio of male-to-hermaphrodite median longevity in the 10 strains was 1.2. In all 10 of the strains, maximum male longevity exceeded that of hermaphrodites (McCulloch and Gems, 2003). All 12 isolates in this study were androdioecious (having males and hermaphrodites) as opposed to dioecious (with separate males and females). To evaluate whether this pattern might be specific to C. elegans or to androdioecious species of free-living soil nematodes like C. elegans, seven other species were examined. Three of these were androdioecious like C. elegans; the others were dioecious. In seven of the eight species, males were the longer-lived sex. In only one species, the androdioecious C. briggsae, were males shorter-lived. Why C. briggsae is exceptional is anything but clear. One aspect of its biology that might (or might not) be relevant is that unlike the other androdioecious species, when C. briggsae males mate with hermaphrodites, the bulk of the progeny are hermaphrodites rather than a 50:50 mix of males and hermaphrodites as in the other species (McCulloch and Gems, 2003). Although the mechanism(s) underlying these sex differences remain obscure, it is provocative that a daf-2 mutation in C. elegans, which doubles longevity in hermaphrodites, more than sextuples it in males (Partridge and Gems, 2002). Understanding these differences is a problem worthy of considerably more investigation.
Fruit Flies
The laboratory fruit fly, Drosophila melanogaster, has likely been subject to more extensive demographic analysis than any other model organism. There have been few analyses of sex differences in longevity, however. Probably the most extensive analysis focused specifically on sex differences is from Malick and Kidwell (1966). They investigated longevity in five isogenic lines and in F1 hybrids among them. Results were complex and highly dependent on genotype, mating status, and female fecundity. Generally, isogenic lines were shorter-lived than hybrids and mating decreased longevity of both sexes. The biggest difference favoring males was a male/female ratio of 3.8 in mean longevity. The biggest difference favoring females was a female/male ratio of 2.3 (Malick and Kidwell, 1966). So sex differences in Drosophila lifespan can be extreme. Similar condition dependence was found in Drosophila subobscura, where unmated females live longer than males, but mating shortens female longevity more, so that mated males outlive mated females (Maynard Smith, 1958). More recently, a study of 219 inbred lines (the Drosophila Genetic Reference Panel) found that in ∼70% of these lines virgin females lived longer than virgin males, although there were lines where virgin males lived more than twice as long as virgin females (Arya et al., 2010). Recent work has identified loss of gut integrity as a major cause of mortality in D. melanogaster (Biteau et al., 2010), and some evidence suggests that sex differences in intestinal stem cell activity may underlie longevity differences in a least some genotypes (Regan et al., 2016).
Interestingly, as with worms, there is also a marked sex difference in the impact of some longevity-enhancing genetic treatments in flies. For instance, reduced signaling in the insulin/IGF pathway enhances longevity substantially in females but has little to no effect on male lifespan (Clancy et al., 2001; Giannakou et al., 2004; Tatar et al., 2001).
While aging in C. elegans and Drosophila has been extensively studied, sex differences in longevity have not been thoroughly investigated. For C. elegans and related species, individual housing leads to longer-lived males compared to hermaphrodites, whereas group housing abolishes this longevity difference. In flies, which sex is longer-lived depends on genotype, mating status, and female fecundity; under some conditions males are much longer-lived and under others females show significantly greater lifespans. Both systems are well characterized and offer many opportunities for investigations into sex-specific mechanisms that influence aging.
Mammals
Females live longer than males in humans and all Old World monkeys and apes for which we have the best data (Austad, 2011; Bronikowski et al., 2011). This appears to be true in both wild and captive populations (Allman et al., 1998; Bronikowski et al., 2011). Yet, whether there is a general mammalian pattern of greater female longevity under protected, captive conditions where intrinsic physiological aging dominates mortality patterns is not known because so few species have been rigorously investigated. What is known about the best-described species indicates that all show a substantially different pattern from humans.
Dogs and Cats
One might expect that there would be loads of data on the relative longevity of male versus female house cats and dogs, the most common species of companion animals in the world. Surprisingly, there is relatively little information on these two domestic species.
The best data on mortality in cats suggest that females have a small survival advantage. Although two early studies found no significant sex differences, one of those was relatively small (Bronson, 1981), and in the other age data were truncated at 13 years, which is about the median age of death in cats (Egenvall et al., 2009). By far the most comprehensive study (of ∼4,000 cats) with complete longevity records, median longevity of females was 2 years or about 15% greater than the longevity of all males (15.0 versus 13.0 years) (O’Neill et al., 2015). In that study as well as in the earlier ones, the impact of neutering on lifespan extension was greater than the impact of sex, however.
For dogs, there is scarcely better information. One small study (n = 287) of laboratory beagles found no significant sex differences in longevity (Albert et al., 1994). However, in a much larger study of thousands of insured Swedish dogs, females were found to be slightly longer-lived (Egenvall et al., 2000). These Swedish results should be interpreted with caution, as no information was available on animals at greater than 10 years of age. Again, the most thorough available data from a study of companion dogs with complete lifespans in the United Kingdom found that after controlling for body weight and pure-breed versus mixed-breed status, males slightly outlived females by 0.4 years among animals surviving to at least 3 years of age (O’Neill et al., 2013). The median longevity in this study was 12.1 years, emphasizing why caution is necessary in interpreting the Swedish data. Consistent with an earlier study (Michell, 1999), O’Neill and colleagues (2013) also found that neutering increased the longevity of females by about 5.6% but did not increase the longevity of males.
In summary, information currently available indicates that in intact cats females are slightly longer-lived and in dogs males are slightly longer-lived. For both species, though, these results should be considered provisional. Also, breed and reproductive status (intact versus neutered) can alter these simple conclusions.
Laboratory Rats
Until the 1990s, laboratory rats were the mammal of choice for most aging studies (Masoro, 1992). Unfortunately, most of those studies focused on males only. However, there have been lifespan studies of laboratory rats of both sexes under a variety of dietary and housing conditions. Most of these studies observed that females live longer than males (Table 1). However, in recent decades two-sex studies have grown rare, so most of what we know about sex differences in rat longevity stems from the time before specific-pathogen-free (SPF) animal colonies became the norm. Thus, it is never clear when infectious disease may have had an undue influence on a study’s outcome. For instance, McCay’s 1935 study of “white” rat longevity observed that female rats lived approximately 66% longer (!) than males when both were fed ad libitum (McCay et al., 1935). However, compared to other rat studies, the males (but not the females) in that study were exceptionally short-lived. Interestingly, when their diets were restricted in calories, the longevity difference was reversed. In two groups restricted either from weaning or 2 weeks after weaning, males lived 6% and 8% longer than females. Most studies since that time have found female rats to live longer than males, although the difference is not dramatic, ranging from a female survival advantage of roughly 2%–15% (Austad, 2011; Sprott and Austad, 1996; Swindell, 2012; Turturro et al., 1999). In particular, the NIA Biomarkers program raised thousands of rats of two inbred strains (F344 and Brown Norway) and one F1 hybrid in both ad libitum-fed and food-restricted conditions, including two different diets, and found that in all cases females lived slightly longer than males. Thus, females appear to have a slight survival advantage in multiple rat genotypes under a variety of conditions.

Table 1
Sex Differences in Laboratory Rat, Rattus norvegicus
Longevity in months. ∗Mean or median, whichever was available. If both were available, mean was used.
Unlike the other model species mentioned so far, there have been few studies of life-extending interventions in rats—none, in fact, except dietary restriction, have been done simultaneously in both sexes. Male response to dietary restriction was greater in 7 of 10 such experiments, including substantial differences in three studies involving the F344 strain alone or hybridized to Brown Norway. The Brown Norway strain by itself had a greater response in females (Sprott and Austad, 1996). These differences are unlikely to be due to chance as they employed sample sizes of approximately 50 animals per sex per group, which is high for rodent studies. However, whether these results are due to the genotype, the details of the diets, or some other idiosyncrasy of husbandry is not clear.
Based on the available data, it appears that female rats are longer-lived than male rats, although the difference is not large; the studies are few and were largely completed prior to the availability of SPF facilities.
Laboratory Mice
There has been a great deal of confusion over sex differences in laboratory mouse longevity (Austad, 2011). One can find statements in the literature that males are the longer-lived sex (Ali et al., 2006), that there is no sex difference (Sanz et al., 2007), or that females live longer (Viña et al., 2005). Each of these claims is correct, it turns out-and wrong.
Wild-derived house mice, those whose ancestors have been in the laboratory for only a few generations, exhibit little sex difference in longevity. For instance, in two of three wild-derived populations from the U.S. mainland and two from Pacific islands all kept under identical conditions in the same animal facility, longevity of the sexes differed by 1% or less. In the third, however, males lived 19% longer than females (Miller et al., 2002).
In order to determine whether this sort of seemingly arbitrary longevity difference was more consistent in a larger sample of studies, Austad (2011) surveyed 118 survival studies of laboratory mice that included either mean or median longevity for both sexes. These were studies that all used standard laboratory conditions without genetic or dietary manipulations, although they could be inbred, F1 hybrid, or outbred populations. The studies included a variety of uncontrolled and unknown husbandry differences including, but certainly not limited to, a range of diets, bedding, housing densities, ambient temperature, and no doubt different pathogen exposure as well. Analysis of these studies clearly showed that mice display no robust or consistent sex differences in longevity. There were substantial sex differences in individual studies though, ranging from males living more than 40% longer to females living more than 70% longer (Storer, 1966). In all, some 65 studies reported that males outlived females, 51 reported that females outlived males, and for the other two studies sex differences were virtually absent.
An obvious suspect to bring some order to this puzzling finding is mouse genetics. All inbred strains of mice or any other species exhibit their own particular idiosyncrasies, of course. Perhaps sex differences in longevity is one particularly common type of strain idiosyncrasy. However, that turns out not to be the case. There were 29 studies that used C57BL/6 mice, and even these studies showed surprising variability. Males lived longer, up to as much as 20% longer, in 18 studies; females lived longer, up to nearly 20% longer, in 11 other studies. Other mouse strains that were assessed in multiple studies also showed longer female lives some times and longer male lives other times.
Because there were so many uncontrolled variables in comparing all these studies, it was not clear whether the range of sex differences observed was due to discoverable environmental differences or to intrinsic stochasticity. Discovering environmental conditions that favor survival of one sex over the other might be informative for understanding the mechanisms underlying sex differences. To investigate this issue, we wished to examine multiple studies of the same mouse genotype when great care had been taken to standardize the husbandry among studies. Fortunately, this is exactly what has been done with the Interventions Testing Program (ITP) (Miller et al., 2007).
The ITP evaluates compounds that can be administered in food or water for their potential ability to extend mouse longevity. There are several unique features of the program, including that it uses genetically heterogeneous mice that are created from the mating of two F1 hybrid lines (CB6F1 × C3D2F1), and performs all experiments at three sites (Michigan, Texas, Maine) simultaneously. The 3-site design makes standardizing procedures between experiments and across sites especially important. Examining the median longevity of control mice from three distinct cohorts at the three sites reveals remarkable consistency, particularly compared to our previous analyses under diverse circumstances (Table 2) (Harrison et al., 2014; Miller et al., 2011; Strong et al., 2008). In all cohorts at all sites, female mice of this genotype lived longer than males. The mean difference was about 13% in median longevity, with 4% being the smallest difference at any site for one experiment and 19% being the largest difference. A fourth cohort also found a 10% difference in the same direction (Strong et al., 2013).
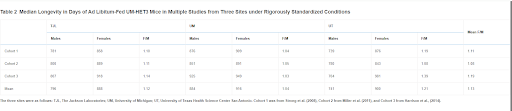
Table 2
Median Longevity in Days of Ad Libitum-Fed UM-HET3 Mice in Multiple Studies from Three Sites under Rigorously Standardized Conditions
The three sites were as follows: TJL, The Jackson Laboratories; UM, University of Michigan; UT, University of Texas Health Science Center San Antonio. Cohort 1 was from Strong et al. (2008), Cohort 2 from Miller et al. (2011), and Cohort 3 from Harrison et al., (2014).
These consistent results from the ITP indicate that the extreme variability observed in the 118 studies may be resolvable into its causal environmental components. If so, then mice can potentially teach us a lot about the underlying mechanisms of sex differences in longevity.
Perhaps even more surprising than the variability in sex differences in mouse longevity across diverse studies is the sex bias found in many interventions in longevity (Table 3) (Austad and Bartke, 2015). This table only includes studies in which a statistically significant effect was found in only one sex and not in the other. There are a multitude of additional studies in which the effect on one sex is substantially greater than the effect in the other, which we have ignored (e.g., Harrison et al., 2009). It is not difficult to imagine why dietary or pharmacological studies might differ between the sexes because of differential metabolism or clearance. This may be the case with NDGA and aspirin, in which blood levels of the active form of the drugs are significantly higher in one sex compared with the other, and could be the case for the other interventions reported to date by the ITP where, with the notable exception of rapamycin, lifespan is preferentially extended in males (Table 4) (Strong et al., 2008). It is more difficult to imagine why something like a genetic knockout—presumably just as knocked out in both sexes—would affect the sexes differently. As with sex differences in unmanipulated mice, there may be a considerable amount to be learned from further investigation of these cases.
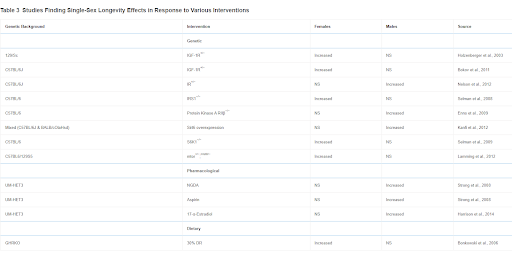
Table 3
Studies Finding Single-Sex Longevity Effects in Response to Various Interventions
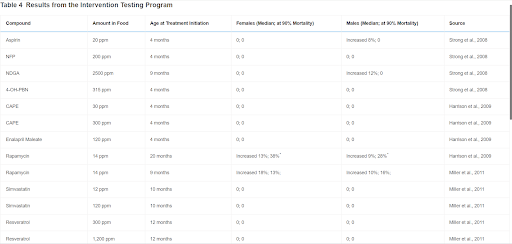
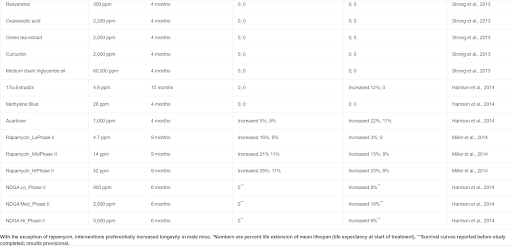
Table 4
Results from the Intervention Testing Program
With the exception of rapamycin, interventions preferentially increased longevity in male mice. ∗Numbers are percent life extension of mean lifespan (life expectancy at start of treatment). ∗∗Survival curves reported before study completed; results provisional.
In summary, there is no consistent sex difference in mouse longevity, although individual studies can be found to support greater male or female longevity. The conditions favoring survival of one sex over the other are also unknown in mice. However, in data from the ITP where conditions and husbandry have been standardized, female median lifespan is consistently great than that of males. These results, coupled with the robust and repeatable sex differences in response to longevity interventions suggest that mice may provide a model for understanding sex differences in aging.
Humans
Consistent sex differences in longevity, robust across diets, mating patterns, and environmental vagaries, have not been seen in any of the species discussed so far. However, in humans, robust sex differences do exist. Indeed, the sex difference in longevity may be one of the most robust features of human biology (Austad, 2011).
Many more orders of magnitude of longevity data are available for humans compared to any other species, but those data are not always reliable. For instance, according to official United Nations data from 1990, residents of Malawi, where life expectancy was only about 40 years, survived better later in life than residents of Japan, the longest-lived country in the world (Austad, 1997). Even with these uncertainties in data quality, however, 176 of the 178 countries, islands, and principalities for which the United Nations currently keeps records have a clear female longevity advantage (http://unstats.un.org/unsd/demographic/products/dyb/dyb2014.htm).
The best source of historical and current human mortality data is the Human Mortality Database (http://www.mortality.org/), which currently has information on 38 countries over periods with particularly reliable data. For instance, complete life tables for both males and females are available from Sweden and France since 1751 and 1816, respectively, but from Japan and Russia only since 1947 and 1959, respectively. Given this high data quality, it is impressive that for all 38 countries for every year in the database, female life expectancy at birth exceeds male life expectancy. The robustness of this pattern is even more apparent when considering that in very early life (birth to age 5) female survival is uniformly better across years and across countries as is later-life survival (life expectancy at age 50) (Austad and Bartke, 2015). Indeed, using data from the Gerontology Research Group (http://www.grg.org/Adams/Tables.htm) to examine sex differences in the oldest of the old, women comprise 90% of supercentenarians (individuals living to 110 years or longer), although the mean age among supercentenarians is comparable between the sexes ( = 111.5 versus 111.3, females and males, respectively).
A particularly vivid example of female survival advantage can be seen in the demographic history of Iceland, a small genetically homogeneous country, historically beset by multiple catastrophes such as famine, flooding, volcanic eruptions, and disease epidemics (Andreeva, 2008). For Icelandic birth cohorts spanning the mid-19th to early 20th century (the latest complete cohort data available), life expectancy at birth fell to as low as 21 years during catastrophes and rose to as high as 69 years during good times. Yet in every year, regardless of food availability or pestilence, women at the beginning of life and near its end survived better than men (Figure 1). Although the sex bias in survival has existed for as long as reliable demographic information is available, it widened for mid- to late-life survival in the very late 19th and early 20th centuries as a consequence of a rapid increase in cardiovascular disease in men, likely due to food superabundance and sex differences in smoking behavior (Beltrán-Sánchez et al., 2015).
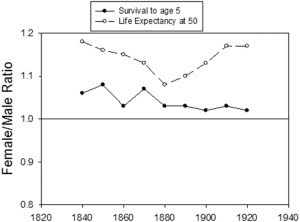
Figure 1 Survival to Age 5 and Life Expectancy at Age 50 in Icelandic Population Cohorts Born between 1840 and 1920
It used to be thought that women’s survival advantage began at conception and was robust in utero as well (Austad, 2011). However, extensive new research on prenatal mortality has now discovered that the advantage begins only several weeks prior to birth (Orzack et al., 2015). Orzack et al. (2015) compiled data on almost 140,000 embryos aged 3–6 days conceived under assisted reproductive technologies (ART) and determined that 50.2% were male. 60% of these embryos had karyotypic abnormalities, consistent with the data reported from inspections of spontaneous abortions. Of those embryos with abnormalities, 50.9% were male, a statistically significant difference, but perhaps not biologically significant. While it can be argued that ART is not the same as natural fertilization, the sex ratio at birth of babies conceived by ART and natural fertilization does not differ (Orzack et al., 2015). Orzack et al. (2015) used additional data from studies of abortion, chorionic villus sampling, and amniocentesis results and determined that male-biased mortality predominated through week 2, followed by female-biased deaths for the next 4 months, succeeded by a period of no sex bias, and finally returning to male-biased mortality during the final 5 weeks prior to birth.
As should be obvious by now, women are not simply resistant to one or two major causes of death. They are less likely to succumb to most of the major causes of death. Of the 15 top causes of death in the United States in 2013, women died at a lower age-adjusted rate of 13 of them, including all of the top 6 causes (Table 5). For one cause, stroke, there was no sex bias, and for one other, Alzheimer’s disease, women were more at risk (Xu et al., 2016).
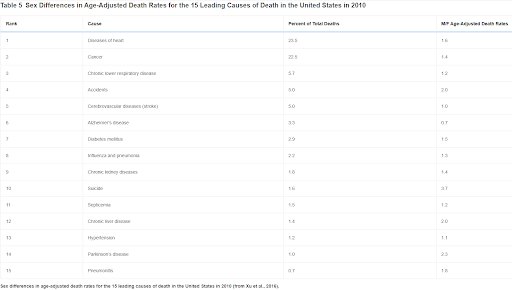
Table 5
Sex Differences in Age-Adjusted Death Rates for the 15 Leading Causes of Death in the United States in 2010
Sex differences in age-adjusted death rates for the 15 leading causes of death in the United States in 2010 (from Xu et al., 2016).
The Mortality-Morbidity Paradox
One of the most puzzling aspects of human sex difference biology, something that has no known equivalent in other species, is that for all their robustness relative to men in terms of survival, women on average appear to be in poorer health than men throughout adult life. This is the mortality-morbidity paradox. In developed Western societies where copious data are available, women make more doctor visits, take more medications, miss more days of work for health reasons, and spend more days in hospitals than men (Christensen et al., 2009; Macintyre et al., 1999; Verbrugge and Wingard, 1987). One possible explanation of this unexpected pattern is that women are more attentive to physical discomfort and illness than men and are more willing to seek medical attention when they experience them (Macintyre et al., 1999). However, the empirical evidence to support this hypothesis is uneven and often contradictory. Moreover, it turns out that the mortality-morbidity paradox is not confined to Western societies, where medical help is easily available, nor is it confined to self-reported data. A higher prevalence of women’s physical limitations in later life has also been reported in Bangladesh, China, Egypt, Guatemala, India, Indonesia, Jamaica, Malaysia, Mexico, the Philippines, Thailand, and Tunisia (Rahman et al., 1994; Wheaton and Crimmins, 2016). Objective measures of sex differences in physical function batteries involving muscle strength, responsiveness, and balance have also shown a female disadvantage in the United States, Taiwan, Korea, China, Indonesia, and even among the Tsimane, an indigenous, non-technological, foraging-horticultural group living in eastern Bolivia (Wheaton and Crimmins, 2016).
What might explain this paradox? There are several plausible hypotheses. The first is that the mortality pattern itself might explain the morbidity pattern (Manton et al., 1995). The so-called mortality-selection hypothesis posits that precisely because men die at higher rates than women throughout life, surviving men are likely to be more physically robust because those who were not would have died. Consistent with this idea are observations such as the fact that men die at a higher age-adjusted rate than women from heart diseases, but morbidity associated with heart disease is greater in women (Wingard et al., 1989). Interestingly, if valid, this hypothesis implies that in any species with a significant sex bias in mortality, we should also observe a mortality-morbidity paradox. The fact that the same pattern has been reported in no other species may reflect that it is a unique human trait or that we do not have the refined data on later-life health in other species that we do in humans. However, in the few studies that have attempted to account for mortality selection statistically, a sex difference in morbidity still remains (Doblhammer and Hoffmann, 2010).
A less-general explanation for the mortality-morbidity paradox is that it has to do with the particular health problems to which humans are prone in later life. It may really be a human-specific phenomenon. Women are well-known to suffer more joint and bone problems, such as osteoarthritis, osteoporosis, and idiopathic back pain, than men (Pinn, 2006). Back and joint pain from arthritis is not only more common, it tends to be more severe among women (Crimmins, 2004; Verbrugge, 1995; Wan et al., 2005). Chronic pain from such conditions not only limits activity, but can also lead sufferers to seek medical help—both features of greater female morbidity—and can also have more far-reaching secondary effects from sequelae such as chronic sleep deprivation and stress. Thus, it is tempting to hypothesize that the sex difference in morbidity is largely due to the more general phenomenon of women’s greater susceptibility to connective tissue maladies. Connective tissue, in humans at least, is known to be particularly responsive to female hormones (Karasik and Ferrari, 2008; Roman-Blas et al., 2009). Sex differences in joint health are not confined to older ages. Women are also considerably more prone to anterior cruciate ligament injury than men during athletic activities (Heitz et al., 1999; Prodromos et al., 2007). To be wildly speculative, perhaps this greater connective tissue responsiveness to hormones may be a side effect of the necessary pelvic ligament responsiveness to hormones during pregnancy and childbirth (Reese and Carey, 2015). In any case, humans are currently the only species in which the mortality-morbidity paradox has been observed.
Mechanisms of Sex Differences in Longevity
Ultimately, of course, we would like to understand the fundamental cellular and molecular mechanisms that account for sex differences in longevity. In every species mentioned above except for humans, sex differences appear to be conditional on some known and some unknown factors. For worms and flies, mating itself may reverse or eliminate sex differences. Genetic factors also affect sex differences as shown by the fact that females live longer in ∼70% of the inbred strains in the Drosophila Genetic Reference Panel, whereas ∼30% of the male strains live longer (Arya et al., 2010). Beyond mating history and genetics, there are also obviously unknown environmental factors at work as well. What, for instance, causes females of a single, widely used mouse genotype, C57BL/6, to live more than 30% longer than males in one study (Cheney et al., 1980) and males to live consistently 10% or more longer in repeated survival studies from another laboratory (Tanaka et al., 2000)?
Although there has been considerable theorizing, there have been relatively few studies specifically addressing mechanisms of sex differences in longevity. These few mechanistic studies have focused most often on humans or laboratory rodents. The two physiological systems most often invoked are the endocrine and immune systems. These of course are known to be related to one another in that sex hormones are well known to affect immune response (Gubbels Bupp, 2015). As was previously noted, lack of mating or sterilization has been repeatedly reported to lengthen life in numerous species, including all species discussed here, suggesting that reproductive hormones may be partially explanatory. In humans in particular, historical records have been examined in Korea indicating that males lacking testosterone, i.e., castrated males, lived 15–20 years longer than intact men of the same socioeconomic strata (Min et al., 2012). By itself, this study would not be particularly compelling as historical records can be mistaken or misleading, but 20th century records on institutionalized men who have been castrated found a surprisingly similar result (Hamilton and Mestler, 1969).
Now that we know that human females survive better than males even in the first few years after birth as well as throughout later life (Austad and Bartke, 2015) (Figure 1), some thought ought to be given to the impact of early-life hormone exposure on both early- and later-life health. In addition to dramatic hormonal differences after puberty, male fetuses exhibit a surge of testosterone during sexual differentiation in utero (Winter et al., 1977), and both sexes experience activation of the hypothalamic-pituitary-gonadal axis in the first few months of postnatal life (Andersson et al., 1998). The increasing realization that early-life events can have manifold later-life health impacts makes the examination of early-life hormonal profiles of particular interest (Bartke et al., 2016; Brakefield et al., 2005; Sonntag et al., 2005). Assuming that the survival advantage is fairly general to female mammals (something that is still unknown), then the anti-oxidant (Mann et al., 2007; Stice et al., 2009) and anti-inflammatory (Benedusi et al., 2012; Villa et al., 2015) properties of estrogen seem like one obvious place to look for an explanation. Both oxidative damage and chronic low-grade inflammatory activation in later life have been implicated in aging (Barja, 2004; Finch, 2007; Ku and Sohal, 1993). Greater steady-state oxidative damage to DNA has been found in peripheral blood leukocytes in men compared to women (Proteggente et al., 2002). Similar sex differences have been found in mtDNA damage from tissues of Wistar rats in which female life expectancy exceeded male life expectancy by 16% (Borras et al., 2007). In this rat study, mitochondria from females were observed to produce more pro-oxidants and have higher anti-oxidant enzyme levels than male mitochondria. The impact of estrogen as a causative agent was verified by ovariectomizing the rats and noting that pro-oxidant levels rose, then fell again when exogenous estrogen was added back.
In addition to oxidative stress, the endocrine impact on immunity and inflammation should not be ignored. Although there have been numerous studies assessing various aspects of immune responsiveness in adult humans in which females have often proven to have more robust responses (Ferguson et al., 2013; Gubbels Bupp, 2015; Oertelt-Prigione, 2012), there have been few such studies in infants. Yet given the early survival differences between the sexes, if differential immune competence could be affecting survival differences, then studies of infants might seem warranted.
Conclusions
A thorough examination of sex differences in longevity in the animal species we know most about has revealed that it is seldom warranted to generalize in claiming that female (or male) worms, flies, dogs, mice, or rats are the longer-lived sex. In virtually every case, it is condition dependent. Thus, in the search for mechanisms underlying sex differences, we need to be aware of, and even take advantage of, the conditions favoring one sex versus the other. In some cases these conditions are known; in others, such as with mice, the conditions remain obscure. However, as single mouse genotypes exhibit greater male longevity sometimes and greater female longevity other times, this offers an excellent system to investigate sex differences further if the predisposing conditions can be identified. The one species with copious data indicating that greater female longevity is virtually universal is humans. The recent discovery that female survival advantage begins at (or perhaps slightly before) birth suggests that investigation of the mechanisms facilitating this early survival difference may yield insight into life-long differences as well.
Acknowledgments
The authors wish to acknowledge support by U.S. NIH/NIA grants P30 AG050886 and R01 AG043972.
References
- Abrams, P.A.
Does increased mortality favor the evolution of more rapid senescence?
Evolution. 1993; 47:877-887
Crossref PubMed Google Scholar
2.Albert, R.E. ∙ Benjamin, S.A. ∙ Shukla, R.
Life span and cancer mortality in the beagle dog and humans
Mech. Ageing Dev. 1994; 74:149-159
Crossref Scopus (40) PubMed Google Scholar
- Ali, S.S. ∙ Xiong, C. ∙ Lucero, J. …
Gender differences in free radical homeostasis during aging: shorter-lived female C57BL6 mice have increased oxidative stress
Aging Cell. 2006; 5:565-574
Crossref Scopus (116) PubMed Google Scholar
- Allman, J. ∙ Rosin, A. ∙ Kumar, R. …
Parenting and survival in anthropoid primates: caretakers live longer
Proc. Natl. Acad. Sci. USA. 1998; 95:6866-6869
Crossref Scopus (112) PubMed Google Scholar
- Andersson, A.M. ∙ Toppari, J. ∙ Haavisto, A.M. …
Longitudinal reproductive hormone profiles in infants: peak of inhibin B levels in infant boys exceeds levels in adult men
- Clin. Endocrinol. Metab. 1998; 83:675-681
Crossref Scopus (366) PubMed Google Scholar
- Andreeva, M. (2008). About mortality data for Iceland. In Human Mortality Database, pp. 1-10.
- Arya, G.H. ∙ Weber, A.L. ∙ Wang, P. …
Natural variation, functional pleiotropy and transcriptional contexts of odorant binding protein genes in Drosophila melanogaster
Genetics. 2010; 186:1475-1485
Crossref Scopus (54) PubMed Google Scholar
- Austad, S.N.
Why We Age
John Wiley & Sons, Inc., New York, 1997
- Austad, S.N.
Why women live longer than men: sex differences in longevity
Gend. Med. 2006; 3:79-92
Abstract Full Text (PDF) Scopus (336) PubMed Google Scholar
- Austad, S.N.
Sex differences in longevity and aging
Masoro, E.J. ∙ Austad, S.N.
The Handbook of the Biology of Aging
Academic Press, San Diego, 2011; 479-496
Crossref Scopus (54) Google Scholar
- Austad, S.N. ∙ Bartke, A.
Sex Differences in Longevity and in Responses to Anti-Aging Interventions: A Mini-Review
Gerontology. 2015; 62:40-46
Crossref Scopus (108) PubMed Google Scholar
- Barja, G.
Free radicals and aging
Trends Neurosci. 2004; 27:595-600
Full Text Full Text (PDF) Scopus (537) PubMed Google Scholar
- Bartke, A. ∙ Sun, L. ∙ Fang, Y. …
Growth hormone actions during development influence adult phenotype and longevity
Exp. Gerontol. 2016; S0531-5565(15)30109-1
- Beltrán-Sánchez, H. ∙ Finch, C.E. ∙ Crimmins, E.M.
Twentieth century surge of excess adult male mortality
Proc. Natl. Acad. Sci. USA. 2015; 112:8993-8998
Crossref Scopus (123) PubMed Google Scholar
- Benedusi, V. ∙ Meda, C. ∙ Della Torre, S. …
A lack of ovarian function increases neuroinflammation in aged mice
Endocrinology. 2012; 153:2777-2788
Crossref Scopus (74) PubMed Google Scholar
- Berg, B.N. ∙ Simms, H.S.
Nutrition and longevity in the rat. II. Longevity and onset of disease with different levels of food intake
- Nutr. 1960; 71:255-263
Crossref Scopus (212) PubMed Google Scholar
- Biteau, B. ∙ Karpac, J. ∙ Supoyo, S. …
Lifespan extension by preserving proliferative homeostasis in Drosophila
PLoS Genet. 2010; 6:e1001159
Crossref Scopus (285) PubMed Google Scholar
- Bokov, A.F. ∙ Garg, N. ∙ Ikeno, Y. …
Does reduced IGF-1R signaling in Igf1r+/- mice alter aging?
PLoS ONE. 2011; 6:e26891
Crossref Scopus (129) PubMed Google Scholar
- Bonkowski, M.S. ∙ Rocha, J.S. ∙ Masternak, M.M. …
Targeted disruption of growth hormone receptor interferes with the beneficial actions of calorie restriction
Proc. Natl. Acad. Sci. USA. 2006; 103:7901-7905
Crossref Scopus (260) PubMed Google Scholar
- Borras, C. ∙ Gambini, J. ∙ Viña, J.
Mitochondrial oxidant generation is involved in determining why females live longer than males
Front. Biosci. 2007; 12:1008-1013
Crossref Scopus (88) PubMed Google Scholar
- Brakefield, P.M. ∙ Gems, D. ∙ Cowen, T. …
What are the effects of maternal and pre-adult environments on ageing in humans, and are there lessons from animal models?
Mech. Ageing Dev. 2005; 126:431-438
Crossref Scopus (41) PubMed Google Scholar
- Bronikowski, A.M. ∙ Altmann, J. ∙ Brockman, D.K. …
Aging in the natural world: comparative data reveal similar mortality patterns across primates
Science. 2011; 331:1325-1328
Crossref Scopus (200) PubMed Google Scholar
- Bronson, R.T.
Age at death of necropsied intact and neutered cats
Am. J. Vet. Res. 1981; 42:1606-1608
- Carey, J.R. ∙ Liedo, P.
Sex mortality differentials and selective survival in large medfly cohorts: implications for human sex mortality differentials
Gerontologist. 1995; 35:588-596
Crossref Scopus (29) PubMed Google Scholar
- Carlson, A.J. ∙ Hoelzel, F.
Apparent prolongation of the life span of rats by intermittent fasting
- Nutr. 1946; 31:363-375
Crossref Scopus (98) PubMed Google Scholar
- Cheney, K.E. ∙ Liu, R.K. ∙ Smith, G.S. …
Survival and disease patterns in C57BL/6J mice subjected to undernutrition
Exp. Gerontol. 1980; 15:237-258
Crossref Scopus (64) PubMed Google Scholar
- Christensen, K. ∙ Doblhammer, G. ∙ Rau, R. …
Ageing populations: the challenges ahead
Lancet. 2009; 374:1196-1208
Full Text Full Text (PDF) Scopus (2680) PubMed Google Scholar
- Clancy, D.J. ∙ Gems, D. ∙ Harshman, L.G. …
Extension of life-span by loss of CHICO, a Drosophila insulin receptor substrate protein
Science. 2001; 292:104-106
Crossref Scopus (1197) PubMed Google Scholar
- Clutton-Brock, T.H.
Reproductive Success
University of Chicago Press, Chicago, 2009
- Clutton-Brock, T.H. ∙ Isvaran, K.
Sex differences in ageing in natural populations of vertebrates
Proc. Biol. Sci. 2007; 274:3097-3104
Crossref Scopus (313) PubMed Google Scholar
- Crimmins, E.M.
Trends in the health of the elderly
Annu. Rev. Public Health. 2004; 25:79-98
Crossref Scopus (346) PubMed Google Scholar
- de Heij, M.E. ∙ van den Hout, P.J. ∙ Tinbergen, J.M.
Fitness cost of incubation in great tits (Parus major) is related to clutch size
Proc. Biol. Sci. 2006; 273:2353-2361
Crossref Scopus (79) PubMed Google Scholar
- Doblhammer, G. ∙ Hoffmann, R.
Gender differences in trajectories of health limitations and subsequent mortality. A study based on the German Socioeconomic Panel 1995-2001 with a mortality follow-up 2002-2005
- Gerontol. B Psychol. Sci. Soc. Sci. 2010; 65:482-491
Crossref Scopus (29) PubMed Google Scholar
- Egenvall, A. ∙ Bonnett, B.N. ∙ Olson, P. …
Gender, age, breed and distribution of morbidity and mortality in insured dogs in Sweden during 1995 and 1996
Vet. Rec. 2000; 146:519-525
Crossref Scopus (79) PubMed Google Scholar
- Egenvall, A. ∙ Nødtvedt, A. ∙ Häggström, J. …
Mortality of life-insured Swedish cats during 1999-2006: age, breed, sex, and diagnosis
- Vet. Intern. Med. 2009; 23:1175-1183
Crossref Scopus (76) PubMed Google Scholar
- Enns, L.C. ∙ Morton, J.F. ∙ Treuting, P.R. …
Disruption of protein kinase A in mice enhances healthy aging
PLoS ONE. 2009; 4:e5963
Crossref Scopus (84) PubMed Google Scholar
- Ferguson, J.F. ∙ Patel, P.N. ∙ Shah, R.Y. …
Race and gender variation in response to evoked inflammation
- Transl. Med. 2013; 11:63
Crossref Scopus (86) PubMed Google Scholar
- Finch, C.E.
Longevity, Senescence, and the Genome
University of Chicago Press, Chicago, 1990
- Finch, C.E.
The biology of human longevity: inflammation, nutrition, and aging in the evolution of lifespans
Academic Press, San Diego, 2007
- Gems, D. ∙ Riddle, D.L.
Genetic, behavioral and environmental determinants of male longevity in Caenorhabditis elegans
Genetics. 2000; 154:1597-1610
Crossref PubMed Google Scholar
- Giannakou, M.E. ∙ Goss, M. ∙ Jünger, M.A. …
Long-lived Drosophila with overexpressed dFOXO in adult fat body
Science. 2004; 305:361
Crossref Scopus (477) PubMed Google Scholar
- Gilbert, C. ∙ Gillman, J. ∙ Loustalot, P. …
The modifying influence of diet and the physical environment of spontaneous tumour frequency in rats
Br. J. Cancer. 1958; 12:565-593
Crossref Scopus (41) PubMed Google Scholar
- Gubbels Bupp, M.R.
Sex, the aging immune system, and chronic disease
Cell. Immunol. 2015; 294:102-110
Crossref Scopus (240) PubMed Google Scholar
- Hamilton, J.B. ∙ Mestler, G.E.
Mortality and survival: comparison of eunuchs with intact men and women in a mentally retarded population
- Gerontol. 1969; 24:395-411
Crossref Scopus (189) PubMed Google Scholar
- Harrison, D.E. ∙ Strong, R. ∙ Sharp, Z.D. …
Rapamycin fed late in life extends lifespan in genetically heterogeneous mice
Nature. 2009; 460:392-395
Crossref Scopus (2985) PubMed Google Scholar
- Harrison, D.E. ∙ Strong, R. ∙ Allison, D.B. …
Acarbose, 17-α-estradiol, and nordihydroguaiaretic acid extend mouse lifespan preferentially in males
Aging Cell. 2014; 13:273-282
Crossref Scopus (293) PubMed Google Scholar
- Heitz, N.A. ∙ Eisenman, P.A. ∙ Beck, C.L. …
Hormonal changes throughout the menstrual cycle and increased anterior cruciate ligament laxity in females
- Athl. Train. 1999; 34:144-149
- Holzenberger, M. ∙ Dupont, J. ∙ Ducos, B. …
IGF-1 receptor regulates lifespan and resistance to oxidative stress in mice
Nature. 2003; 421:182-187
Crossref Scopus (1707) PubMed Google Scholar
- Iliadi, K.G. ∙ Iliadi, N.N. ∙ Boulianne, G.L.
Regulation of Drosophila life-span: effect of genetic background, sex, mating and social status
Exp. Gerontol. 2009; 44:546-553
Crossref Scopus (26) PubMed Google Scholar
- Kanfi, Y. ∙ Naiman, S. ∙ Amir, G. …
The sirtuin SIRT6 regulates lifespan in male mice
Nature. 2012; 483:218-221
Crossref Scopus (875) PubMed Google Scholar
51.Karasik, D. ∙ Ferrari, S.L.
Contribution of gender-specific genetic factors to osteoporosis risk
Ann. Hum. Genet. 2008; 72:696-714
Crossref Scopus (59) PubMed Google Scholar
- Kasuya, T. ∙ Marsh, H.
Life history and reproductive biology of the short-finned pilot whalte, Globicephala macrorhynchus
Rep. Int. Whaling Comm. 1984; 6:259-310
- Ku, H.H. ∙ Sohal, R.S.
Comparison of mitochondrial pro-oxidant generation and anti-oxidant defenses between rat and pigeon: possible basis of variation in longevity and metabolic potential
Mech. Ageing Dev. 1993; 72:67-76
Crossref Scopus (207) PubMed Google Scholar
- Lamming, D.W. ∙ Ye, L. ∙ Katajisto, P. …
Rapamycin-induced insulin resistance is mediated by mTORC2 loss and uncoupled from longevity
Science. 2012; 335:1638-1643
Crossref Scopus (974) PubMed Google Scholar
- Leuze, C.C.K.
The application of radiotracking and its effect on the behavioral ecology of the water vole
Amlamer, D.W.
A Handbook on Biotelemetry and Radio Tracking. C.J.M
Pergamon, Oxford, 1980; 361-366
- Liker, A. ∙ Székely, T.
Mortality costs of sexual selection and parental care in natural populations of birds
Evolution. 2005; 59:890-897
Crossref Scopus (205) PubMed Google Scholar
- Macintyre, S. ∙ Ford, G. ∙ Hunt, K.
Do women ‘over-report’ morbidity? Men’s and women’s responses to structured prompting on a standard question on long standing illness
Soc. Sci. Med. 1999; 48:89-98
Crossref Scopus (202) PubMed Google Scholar
- Malick, L.E. ∙ Kidwell, J.F.
The effect of mating status, sex and genotype on longevity in Drosophila melanogaster
Genetics. 1966; 54:203-209
- Mann, V. ∙ Huber, C. ∙ Kogianni, G. …
The antioxidant effect of estrogen and Selective Estrogen Receptor Modulators in the inhibition of osteocyte apoptosis in vitro
Bone. 2007; 40:674-684
Full Text Full Text (PDF) Scopus (104) PubMed Google Scholar
- Manton, K.G. ∙ Woodbury, M.A. ∙ Stallard, E.
Sex differences in human mortality and aging at late ages: the effect of mortality selection and state dynamics
Gerontologist. 1995; 35:597-608
Crossref Scopus (23) PubMed Google Scholar
- Masoro, E.J.
The role of animal models in meeting the gerontologic challenge of the 21st century
Gerontologist. 1992; 32:627-633
Crossref Scopus (15) PubMed Google Scholar
- Maynard Smith, J.
Prolongation of life in Drosophila subobscura
Proc. R. Soc. Lond. B Biol. Sci. 1958; 144:159-171
- McCay, C.M. ∙ Crowell, M.F. ∙ Maynard, L.A.
The effect of retarded growth upon the length of the life span and upon ultimate body size
J.Nutr. 1935; 13:669-679
- McCulloch, D. ∙ Gems, D.
Evolution of male longevity bias in nematodes
Aging Cell. 2003; 2:165-173
Crossref Scopus (61) PubMed Google Scholar
- Michell, A.R.
Longevity of British breeds of dog and its relationships with sex, size, cardiovascular variables and disease
Vet. Rec. 1999; 145:625-629
Crossref Scopus (213) PubMed Google Scholar
- Miller, R.A. ∙ Harper, J.M. ∙ Dysko, R.C. …
Longer life spans and delayed maturation in wild-derived mice
Exp. Biol. Med. (Maywood). 2002; 227:500-508
Crossref Scopus (196) PubMed Google Scholar
- Miller, R.A. ∙ Harrison, D.E. ∙ Astle, C.M. …
An Aging Interventions Testing Program: study design and interim report
Aging Cell. 2007; 6:565-575
Crossref Scopus (165) PubMed Google Scholar
- Miller, R.A. ∙ Harrison, D.E. ∙ Astle, C.M. …
Rapamycin, but not resveratrol or simvastatin, extends life span of genetically heterogeneous mice
- Gerontol. A Biol. Sci. Med. Sci. 2011; 66:191-201
Crossref Scopus (746) PubMed Google Scholar
- Miller, R.A. ∙ Harrison, D.E. ∙ Astle, C.M. …
Rapamycin-mediated lifespan increase in mice is dose and sex dependent and metabolically distinct from dietary restriction
Aging Cell. 2014; 13:468-477
Crossref Scopus (450) PubMed Google Scholar
- Min, K.J. ∙ Lee, C.K. ∙ Park, H.N.
The lifespan of Korean eunuchs
Curr. Biol. 2012; 22:R792-R793
Full Text Full Text (PDF) Scopus (143) PubMed Google Scholar
- Moorad, J.A. ∙ Promislow, D.E.
Evolution: aging up a tree?
Curr. Biol. 2010; 20:R406-R408
Full Text Full Text (PDF) Scopus (23) PubMed Google Scholar
- Nelson, J.F. ∙ Strong, R. ∙ Bokov, A. …
Probing the relationship between insulin sensitivity and longevity using genetically modified mice
- Gerontol. A Biol. Sci. Med. Sci. 2012; 67:1332-1338
Crossref Scopus (29) PubMed Google Scholar
- Nolan, G.A.
Effect of various restricted dietary regimens on growth, health and longevity of albino rats
- Nutr. 1972; 102:1477-1493
- Nussey, D.H. ∙ Kruuk, L.E. ∙ Morris, A. …
Inter- and intrasexual variation in aging patterns across reproductive traits in a wild red deer population
Am. Nat. 2009; 174:342-357
Crossref Scopus (143) PubMed Google Scholar
- O’Neill, D.G. ∙ Church, D.B. ∙ McGreevy, P.D. …
Longevity and mortality of owned dogs in England
Vet. J. 2013; 198:638-643
Crossref Scopus (218) PubMed Google Scholar
- O’Neill, D.G. ∙ Church, D.B. ∙ McGreevy, P.D. …
Longevity and mortality of cats attending primary care veterinary practices in England
- Feline Med. Surg. 2015; 17:125-133
Crossref Scopus (151) PubMed Google Scholar
- Oertelt-Prigione, S.
The influence of sex and gender on the immune response
Autoimmun. Rev. 2012; 11:A479-A485
Crossref Scopus (349) PubMed Google Scholar
- Orzack, S.H. ∙ Stubblefield, J.W. ∙ Akmaev, V.R. …
The human sex ratio from conception to birth
Proc. Natl. Acad. Sci. USA. 2015; 112:E2102-E2111
Crossref Scopus (181) PubMed Google Scholar
- Partridge, L. ∙ Gems, D.
Mechanisms of ageing: public or private?
Nat. Rev. Genet. 2002; 3:165-175
Crossref Scopus (414) PubMed Google Scholar
- Pinn, V.W.
Past and future: sex and gender in health research, the aging experience, and implications for musculoskeletal health
Orthop. Clin. North Am. 2006; 37:513-521
Full Text Full Text (PDF) Scopus (9) PubMed Google Scholar
- Podlutsky, A.J. ∙ Khritankov, A.M. ∙ Ovodov, N.D. …
A new field record for bat longevity
- Gerontol. A Biol. Sci. Med. Sci. 2005; 60:1366-1368
Crossref Scopus (112) PubMed Google Scholar
- Prodromos, C.C. ∙ Han, Y. ∙ Rogowski, J. …
A meta-analysis of the incidence of anterior cruciate ligament tears as a function of gender, sport, and a knee injury-reduction regimen
Arthroscopy. 2007; 23:1320-1325.e6, e1326
Full Text Full Text (PDF) Scopus (685) PubMed Google Scholar
83.Promislow, D.E.L.
Costs of sexual selection in natural populations of mammals
Proc.Roy.Soc.Lond. Bol. Biol. 1992; 247:203-210
Crossref Scopus (259) Google Scholar
- Promislow, D.E.L. ∙ Montgomerie, R. ∙ Martin, T.E.
Mortality costs of sexual dimorphism in birds
Proc. R. Soc. Lond. B. 1992; 250:143-150
Crossref Scopus (245) Google Scholar
- Proteggente, A.R. ∙ England, T.G. ∙ Rehman, A. …
Gender differences in steady-state levels of oxidative damage to DNA in healthy individuals
Free Radic. Res. 2002; 36:157-162
Crossref Scopus (51) PubMed Google Scholar
- Rahman, O. ∙ Strauss, J. ∙ Gertler, P. …
Gender differences in adult health: an international comparison
Gerontologist. 1994; 34:463-469
Crossref Scopus (55) PubMed Google Scholar
- Reese, M.E. ∙ Carey, E.
Hormonal influence on the neuromusculoskeletal system in pregnancy and postpartum
Fitzgerald, C.M. ∙ Segal, N.A.
Musculoskeletal Health in Pregnancy
Springer International, Switzerland, 2015; 19-39
- Regan, J.C. ∙ Khericha, M. ∙ Dobson, A.J. …
Sex difference in pathology of the ageing gut mediates the greater response of female lifespan to dietary restriction
eLife. 2016; 5:5
Crossref Scopus (158) Google Scholar
- Roman-Blas, J.A. ∙ Castañeda, S. ∙ Largo, R. …
Osteoarthritis associated with estrogen deficiency
Arthritis Res. Ther. 2009; 11:241
Crossref Scopus (245) PubMed Google Scholar
- Sanz, A. ∙ Hiona, A. ∙ Kujoth, G.C. …
Evaluation of sex differences on mitochondrial bioenergetics and apoptosis in mice
Exp. Gerontol. 2007; 42:173-182
Crossref Scopus (59) PubMed Google Scholar
- Selman, C. ∙ Lingard, S. ∙ Choudhury, A.I. …
Evidence for lifespan extension and delayed age-related biomarkers in insulin receptor substrate 1 null mice
FASEB J. 2008; 22:807-818
Crossref Scopus (437) PubMed Google Scholar
- Selman, C. ∙ Tullet, J.M. ∙ Wieser, D. …
Ribosomal protein S6 kinase 1 signaling regulates mammalian life span
Science. 2009; 326:140-144
Crossref Scopus (956) PubMed Google Scholar
93.Sonntag, W.E. ∙ Carter, C.S. ∙ Ikeno, Y. …
Adult-onset growth hormone and insulin-like growth factor I deficiency reduces neoplastic disease, modifies age-related pathology, and increases life span
Endocrinology. 2005; 146:2920-2932
Crossref Scopus (124) PubMed Google Scholar
- Sprott, R.L. ∙ Austad, S.N.
Animal models for Aging Research
Schneider, E.L. ∙ Rowe, J.W.
Handbook of the Biology of Aging
Academic Press, San Diego, 1996; 3-23
- Stice, J.P. ∙ Lee, J.S. ∙ Pechenino, A.S. …
Estrogen, aging and the cardiovascular system
Future Cardiol. 2009; 5:93-103
Crossref Scopus (49) PubMed Google Scholar
- Storer, J.B.
Longevity and gross pathology at death in 22 inbred mouse strains
- Gerontol. 1966; 21:404-409
Crossref Scopus (189) PubMed Google Scholar
- Strong, R. ∙ Miller, R.A. ∙ Astle, C.M. …
Nordihydroguaiaretic acid and aspirin increase lifespan of genetically heterogeneous male mice
Aging Cell. 2008; 7:641-650
Crossref Scopus (259) PubMed Google Scholar
- Strong, R. ∙ Miller, R.A. ∙ Astle, C.M. …
Evaluation of resveratrol, green tea extract, curcumin, oxaloacetic acid, and medium-chain triglyceride oil on life span of genetically heterogeneous mice
- Gerontol. A Biol. Sci. Med. Sci. 2013; 68:6-16
Crossref Scopus (178) PubMed Google Scholar
- Swindell, W.R.
Dietary restriction in rats and mice: a meta-analysis and review of the evidence for genotype-dependent effects on lifespan
Ageing Res. Rev. 2012; 11:254-270
Crossref Scopus (173) PubMed Google Scholar
- Tanaka, S. ∙ Segawa, T. ∙ Tamaya, N. …
Establishment of an Aging Farm of F344/N Rats and C57BL/6 Mice at the National Institute for Longevity Sciences (NILS)
Arch. Gerontol. Geriatr. 2000; 30:215-223
Full Text Full Text (PDF) Scopus (41) PubMed Google Scholar
- Tatar, M. ∙ Kopelman, A. ∙ Epstein, D. …
A mutant Drosophila insulin receptor homolog that extends life-span and impairs neuroendocrine function
Science. 2001; 292:107-110
Crossref Scopus (1320) PubMed Google Scholar
- Tower, J.
Sex-specific regulation of aging and apoptosis
Mech. Ageing Dev. 2006; 127:705-718
Crossref Scopus (110) PubMed Google Scholar
- Trivers, R.L.
Parental investment and sexual selection
Campbell, B.
Sexual selection and the descent of man
Aldine-Atherton, Chicago, IL, 1972; 136-179
- Trivers, R.L.
Social evolution
Benjamin-Cummings Publ. Co., Menlo Park, Calif, 1985
- Turturro, A. ∙ Witt, W.W. ∙ Lewis, S. …
Growth curves and survival characteristics of the animals used in the Biomarkers of Aging Program
- Gerontol. A Biol. Sci. Med. Sci. 1999; 54:B492-B501
Crossref Scopus (557) PubMed Google Scholar
- Verbrugge, L.M.
Women, men, and osteoarthritis
Arthritis Care Res. 1995; 8:212-220
Crossref Scopus (79) PubMed Google Scholar
- Verbrugge, L.M. ∙ Wingard, D.L.
Sex differentials in health and mortality
Women Health. 1987; 12:103-145
Crossref Scopus (253) PubMed Google Scholar
- Villa, A. ∙ Rizzi, N. ∙ Vegeto, E. …
Estrogen accelerates the resolution of inflammation in macrophagic cells
Sci. Rep. 2015; 5:15224
Crossref Scopus (175) PubMed Google Scholar
- Viña, J. ∙ Borrás, C. ∙ Gambini, J. …
Why females live longer than males: control of longevity by sex hormones
Sci. SAGE KE. 2005; 2005:pe17
- Wan, H., Sengupta, M., Velkoff, V.A., and DeBarros, K.A. (2005). 65+ in the United States: 2005. In Current Population Reports, pp. 23-209.
- Waser, P.M. ∙ Jones, W.T.
Survival and reproductive effort in banner-tailed kangaroo rats
Ecology. 1991; 72:771-777
Crossref Scopus (37) Google Scholar
- Wheaton, F.V. ∙ Crimmins, E.M.
Female disability disadvantage: a global perspective on sex differences in physical function and disability
Ageing Soc. 2016; Published online May 8, 2015
Crossref Scopus (53) PubMed Google Scholar
- Williams, G.C.
Pleiotropy, natural selection, and the evolution of senescence
Evolution. 1957; 11:398-411
- Wingard, D.L. ∙ Cohn, B.A. ∙ Kaplan, G.A. …
Sex differentials in morbidity and mortality risks examined by age and cause in the same cohort
Am. J. Epidemiol. 1989; 130:601-610
Crossref Scopus (80) PubMed Google Scholar
- Winter, J.S. ∙ Faiman, C. ∙ Reyes, F.I.
Sex steroid production by the human fetus: its role in morphogenesis and control by gonadotropins
Birth Defects Orig. Artic. Ser. 1977; 13:41-58
- Xu, J. ∙ Murphy, S.L. ∙ Kochanek, K.D. …
Deaths: Final Data for 2013
Natl. Vital Stat. Rep. 2016; 64:1-119
Soure: Cell.com
Read more: